PUBLICADO POR: MARIA DEL REFUGIO DENISE CLAVIJO CORNEJO
UAM IZTAPALAPA
BIOLOGÍA CELULAR
NORMA EDITH LÓPEZ DIAZ-GUERRERO
BIOLOGÍA CELULAR
NORMA EDITH LÓPEZ DIAZ-GUERRERO
 La membrana celular define la periferia de las células, separando su contenido del medio exterior. Esta compuesto de lípidos y proteínas que forman una barrera hidrofobita alrededor de la célula.
La membrana celular define la periferia de las células, separando su contenido del medio exterior. Esta compuesto de lípidos y proteínas que forman una barrera hidrofobita alrededor de la célula.

 Los fosfolípidos son los lípidos más abundantes en las membranas, debido a su carácter anfipático, y en un medio acuoso se organizan espontáneamente conformando la denominada bicapa lipídica. Las cabezas polares están colocadas hacia el medio acuoso (intra y extracelular) y las colas hidrofóbicas hacia el medio lipídico, es decir, al interior de la bicapa, constituyendo la matriz de la membrana. A su vez, estas bicapas tienden a cerrarse espontáneamente sobre sí mismas formando vesículas, es decir, compartimientos cerrados en toda su extensión tridimensional, similares a una esfera.
Los fosfolípidos son los lípidos más abundantes en las membranas, debido a su carácter anfipático, y en un medio acuoso se organizan espontáneamente conformando la denominada bicapa lipídica. Las cabezas polares están colocadas hacia el medio acuoso (intra y extracelular) y las colas hidrofóbicas hacia el medio lipídico, es decir, al interior de la bicapa, constituyendo la matriz de la membrana. A su vez, estas bicapas tienden a cerrarse espontáneamente sobre sí mismas formando vesículas, es decir, compartimientos cerrados en toda su extensión tridimensional, similares a una esfera. Las cadenas hidrocarbonadas de los ácidos grasos que forman parte los fosfolípidos (también denominadas “colas” o grupos acilo), pueden presentarse:
Las cadenas hidrocarbonadas de los ácidos grasos que forman parte los fosfolípidos (también denominadas “colas” o grupos acilo), pueden presentarse: Las funciones del colesterol se pueden resumir de la siguiente manera
Las funciones del colesterol se pueden resumir de la siguiente manera Factores que aumentan la fluidez de las membranas
Factores que aumentan la fluidez de las membranas

 En ambas caras de la bicapa no se encuentran los mismos tipos de fosfolípidos. Si bien estos en su mayoría se sintetizan en la cara citosolica del retículo endoplasmático liso, luego, por medio de movimientos del tipo flip-flop (únicamente permitidos en el REL, gracias a la presencia de flipasas), se van ubicando del lado de la bicapa que les corresponda. Por ej., la fosfatidilcolina y la esfingomielina predominan en la cara no citosolica de la membrana.
En ambas caras de la bicapa no se encuentran los mismos tipos de fosfolípidos. Si bien estos en su mayoría se sintetizan en la cara citosolica del retículo endoplasmático liso, luego, por medio de movimientos del tipo flip-flop (únicamente permitidos en el REL, gracias a la presencia de flipasas), se van ubicando del lado de la bicapa que les corresponda. Por ej., la fosfatidilcolina y la esfingomielina predominan en la cara no citosolica de la membrana. Flopasas. La segunda clase de transportadores de lípidos que dependen de ATP son las flopasas que están dirigidas hacia la cara externa. Los transportadores ABC son un grupo diverso de proteínas que en general son responsables de la exportación dependiente de ATP de los componentes amfipáticos. Algunos miembros de estos transportadores demuestran especificidad para sus substratos respectivos. Las actividades de las flopasa lipídicas mejor caracterizadas son aquellas catalizadas por ABCA1, ABCB1, ABCB4 Y ABCC1.
Flopasas. La segunda clase de transportadores de lípidos que dependen de ATP son las flopasas que están dirigidas hacia la cara externa. Los transportadores ABC son un grupo diverso de proteínas que en general son responsables de la exportación dependiente de ATP de los componentes amfipáticos. Algunos miembros de estos transportadores demuestran especificidad para sus substratos respectivos. Las actividades de las flopasa lipídicas mejor caracterizadas son aquellas catalizadas por ABCA1, ABCB1, ABCB4 Y ABCC1. Sin embargo no toda la membrana tiene la misma fluidez, existen balsas lipidicas o sectores que tiene una fluidez distinta llamados microdominios de membranas que tienen una composición lipidica mas rica en colesterol por lo tanto son menos fluidos.
Sin embargo no toda la membrana tiene la misma fluidez, existen balsas lipidicas o sectores que tiene una fluidez distinta llamados microdominios de membranas que tienen una composición lipidica mas rica en colesterol por lo tanto son menos fluidos.
 Es el mecanismo de pasaje para sustancias que en general son insolubles en lípidos, de tamaños moleculares mayores, iones o moléculas polares que no pueden atravesar el interior hidrofóbico. estas sustancias se combinan con proteínas integrales específicas de la membrana plasmática. Se realiza a favor de un gradiente de concentración, sin gasto de energía.
Es el mecanismo de pasaje para sustancias que en general son insolubles en lípidos, de tamaños moleculares mayores, iones o moléculas polares que no pueden atravesar el interior hidrofóbico. estas sustancias se combinan con proteínas integrales específicas de la membrana plasmática. Se realiza a favor de un gradiente de concentración, sin gasto de energía.  • Transporte de Masa:
• Transporte de Masa: Los desmosomas son estructuras celulares que mantienen adheridas a células vecinas. Estructuralmente dicha unión está mediada por cadherinas (desmogleína y desmocolina), a sus filamentos intermedios (queratina). En el interior de las células actúan como lugares de anclaje para los filamentos intermedios en forma de cuerda, los cuales forman una red estructural en el citoplasma proporcionando una cierta rigidez. Mediante estas uniones los filamentos intermedios de las células adyacentes están indirectamente conectados formando una red continua que se extiende a todo el tejido. El tipo de filamentos intermedios anclados a los desmosomas depende del tipo celular: de queratina en la mayoría de las células epiteliales y de desmina en las fibras musculares cardíacas.
Los desmosomas son estructuras celulares que mantienen adheridas a células vecinas. Estructuralmente dicha unión está mediada por cadherinas (desmogleína y desmocolina), a sus filamentos intermedios (queratina). En el interior de las células actúan como lugares de anclaje para los filamentos intermedios en forma de cuerda, los cuales forman una red estructural en el citoplasma proporcionando una cierta rigidez. Mediante estas uniones los filamentos intermedios de las células adyacentes están indirectamente conectados formando una red continua que se extiende a todo el tejido. El tipo de filamentos intermedios anclados a los desmosomas depende del tipo celular: de queratina en la mayoría de las células epiteliales y de desmina en las fibras musculares cardíacas.

 La matriz extracelular (MEC) es una entidad estructuralmente compleja que rodea y soporta las células que se encuentran en los tejidos de los mamíferos. La MEC también es comúnmente conocida como tejido conectivo.
La matriz extracelular (MEC) es una entidad estructuralmente compleja que rodea y soporta las células que se encuentran en los tejidos de los mamíferos. La MEC también es comúnmente conocida como tejido conectivo.

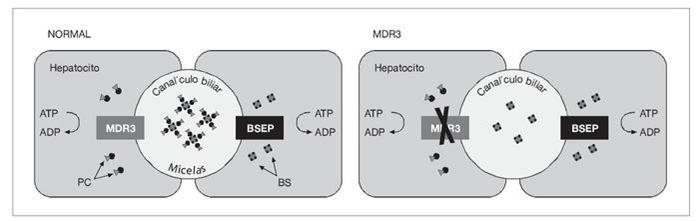
 B. Uniones celulares
B. Uniones celulares 

{kind=link}
{kind=link}
{kind=link}